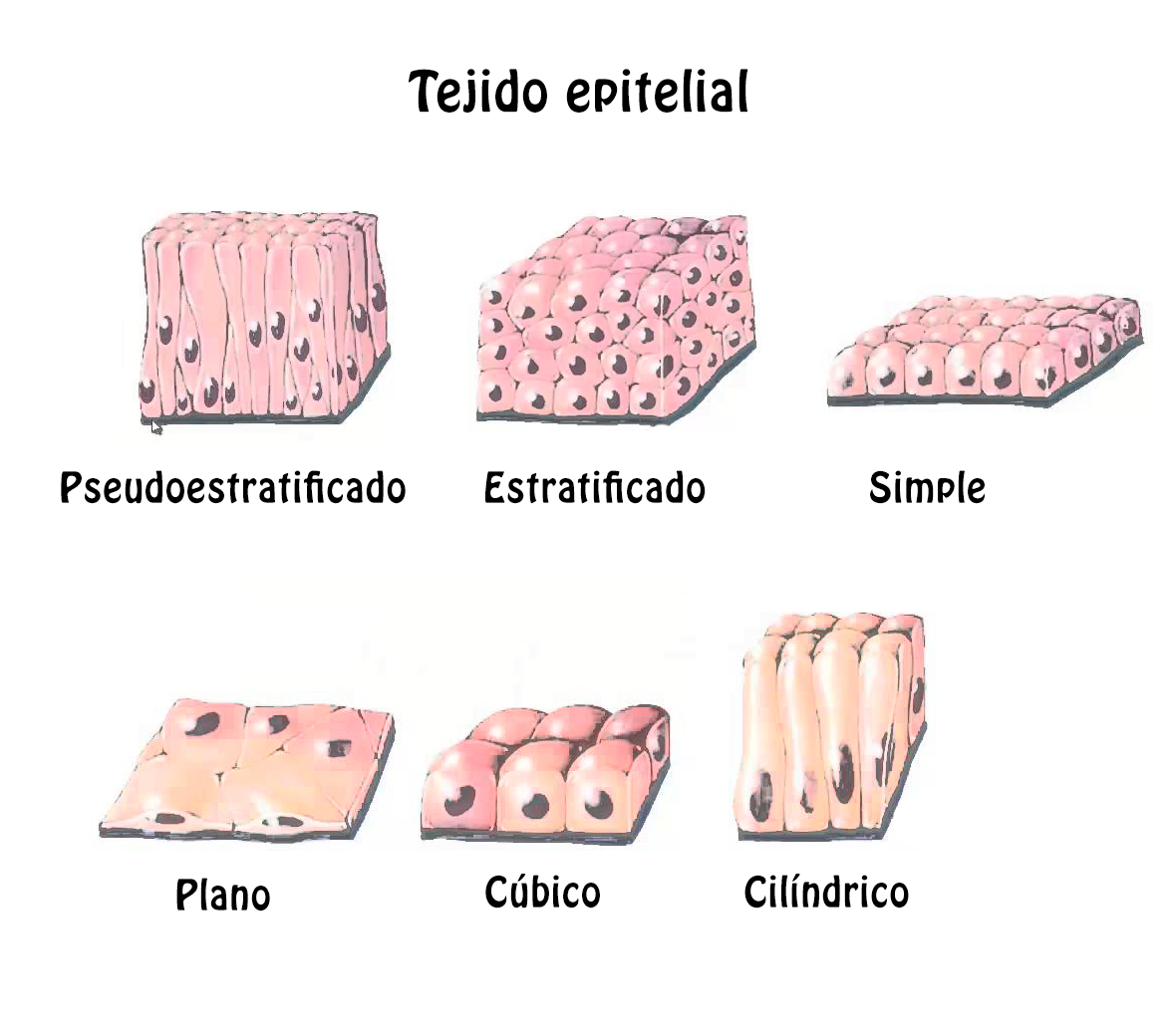
Todas las superficies externas del cuerpo, así como las de las cavidades, conductos y sacos internos, se encuentran recubiertas por células a partir de la membrana basal del ectodermo y el endodermo, conocidas de manera general como células epiteliales, epitelio o tejido epitelial. Este tejido se integra con una o varias capas de células y es avascular (no contiene vasos sanguíneos), pero crece sobre un tejido conectivo subyacente rico en vasos, al cual se une mediante una capa de sostén extracelular denominada membrana basal.

Con frecuencia, el tejido conectivo forma evaginaciones denominadas papilas.
Durante el desarrollo embrionario, los epitelios que recubren las superficies pueden formar invaginaciones en el tejido conectivo subyacente y formar glándulas. En consecuencia, éstas conforman el parénquima (porción secretora) y sus conductos excretores. Por otra parte, también hay células epiteliales especializadas que funcionan como receptores sensoriales (visión, oído, gusto y olfato).
Las funciones del epitelio son variadas. La superficie libre protege contra el estrés mecánico, el acceso de microorganismos, la pérdida de agua y, por otro lado, tiene una participación importante en el sentido del tacto. En las superficies internas su función, en la mayor parte de los casos, es de absorción o secreción y, en otros casos, funciona tan sólo como barrera de protección.
Las células epiteliales poseen tres características: a) se adhieren entre sí por medio de uniones intercelulares especializadas; estas estructuras relacionadas con la membrana plasmática contribuyen a la adherencia y comunicación entre las células; b) la región basal posee una superficie de contacto entre las células epiteliales y el tejido conectivo subyacente llamada membrana basal, y c) tienen una región apical, una región lateral y una región basal, cada una de ellas con funciones diferentes.

ECTODERMO:
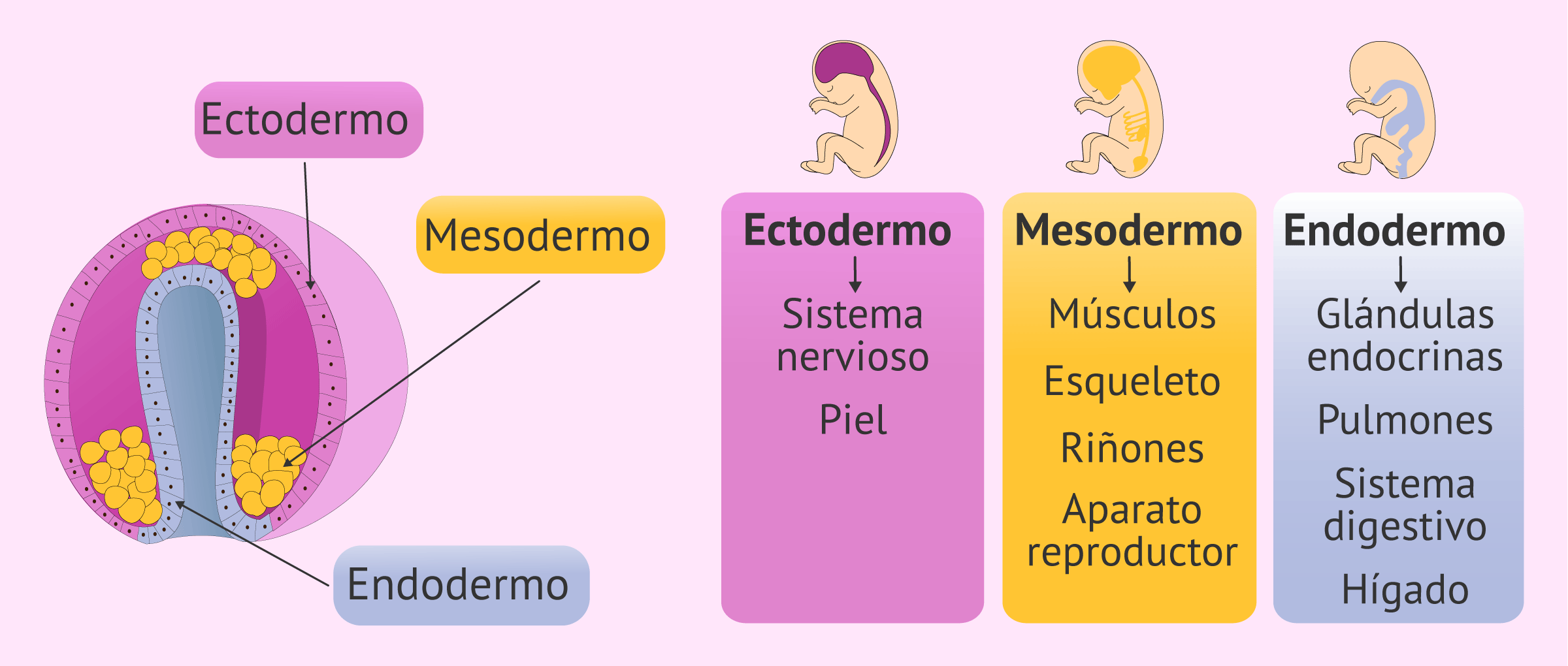
El ectodermo es la capa más exterior del embrión animal durante sus primeras fases de desarrollo. Tras la fecundación en la reproducción de los seres vivos la célula inicial, llamada cigoto, se empieza a dividir y pasa por los estadios de mórula y blástula, a medida que el número de células del embrión aumenta.
En un momento determinado del desarrollo el número celular es muy alto y el embrión toma la forma de una esfera hueca con el interior relleno de líquido pero sin presencia celular; a este estadio se le denomina ya ectodermo. Sin embargo durante la evolución de los seres vivos el sistema embrionario de divisiones se ha ido volviendo cada vez más complejo, para dar lugar a la gran complejidad de organismos existentes.
A partir del ectodermo se formarán, mediante la invaginación de una sección determinada de las células de la esfera, el endodermo y el mesodermo.
El ectodermo da lugar por norma general a tejidos epiteliales, como la piel y todas las estructuras accesorias que colaboran con este órgano. Entre ellos el ectodermo forma el pelo, las plumas, los cuernos, uñas, pezuñas y todas las glándulas que colaboran, sebáceas e incluso las mamarias. Además forma el estomodeo, una invaginación del ectodermo que dará lugar a la boca y el epitelio que la recubre (el tubo digestivo superior) y el interior de la nariz (denominados en conjunto como las vías respiratorias altas). Estos tejidos quedan separados de la faringe, de origen endodérmico, por una membrana que desaparece al día 24 de vida intrauterina.
Además del ectodermo derivan en la cresta neural y en el tubo neural en los cordados. Estos se forman por una invaginación secundaria del ectodermo. De la cresta y el tubo neural se desarrollarán todo el sistema nervioso; el sistema nervioso periférico de la cresta neural y el cerebro, los nervios motores y la médula espinal a partir del tubo neural.


Partes del ectodermo
Ectodermo externo o superficial
Esta zona es la que da lugar a los tejidos epiteliales como las glándulas de la piel, la boca, las cavidades nasales, el pelo, las uñas, parte de los ojos, etc. En animales, origina plumas, cuernos y pezuñas.
Cresta neural
Existen cuatro vías migratorias diferentes para la diferenciación de las células de la cresta neural. Cada vía determina en qué estructuras celulares específicas van a transformarse. Así, van a dar lugar a:
Cresta neural
Existen cuatro vías migratorias diferentes para la diferenciación de las células de la cresta neural. Cada vía determina en qué estructuras celulares específicas van a transformarse. Así, van a dar lugar a:
– Las neuronas y células gliales de los ganglios sensitivos, que son componentes fundamentales del sistema nervioso periférico.
– Las neuronas y glía de los ganglios autónomos, que incluyen los ganglios del sistema nervioso simpático y parasimpático.
– Células neurosecretoras de las glándulas suprarrenales, que se incluyen en la parte dorsal de los riñones.
– Células que van a transformarse en tejidos no neurales, como los melanocitos. Éstos últimos tienen el objetivo de producir la melanina de la piel. También hay grupos de células que van a conformar el cartílago de la cara y los dientes.
Tubo neural
El tubo neural se cierra como si fuera una cremallera. Comienza en la región cervical, y desde ahí prosigue en dirección craneal y caudal. Hasta que la fusión finalice, los extremos craneal y caudal del tubo neural permanecen abiertos, comunicados con la cavidad amniótica.
Cuando se cierra el extremo más craneal, aparecen unas dilataciones llamadas vesículas encefálicas. Éstas son las que van a dar lugar al encéfalo, concretamente a sus primeras divisiones: el romboencéfalo, el mesencéfalo y el prosencéfalo.
Por otra parte, las células del tubo neural también van a constituir la retina de los ojos y la neurohipófisis. Éste último es el lóbulo posterior de la glándula pituitaria.Las dos últimas partes reciben el nombre de neuroectodermo.
Partes del cuerpo derivadas del ectodermo
El ectodermo deriva en las siguientes estructuras:
– Sistema nervioso (cerebro, médula espinal y nervios periféricos).
– Epidermis.
– Glándulas sudoríparas y mamarias.
– Esmalte dental.
– Revestimiento de la boca, fosas nasales y ano.
– Pelo y uñas.
– Los cristalinos de los ojos.
– Partes de oído interno.
VIDEO COMPLEMENTARIO:
https://www.youtube.com/watch?v=fCmdjx8RUDk
MESODERMO:
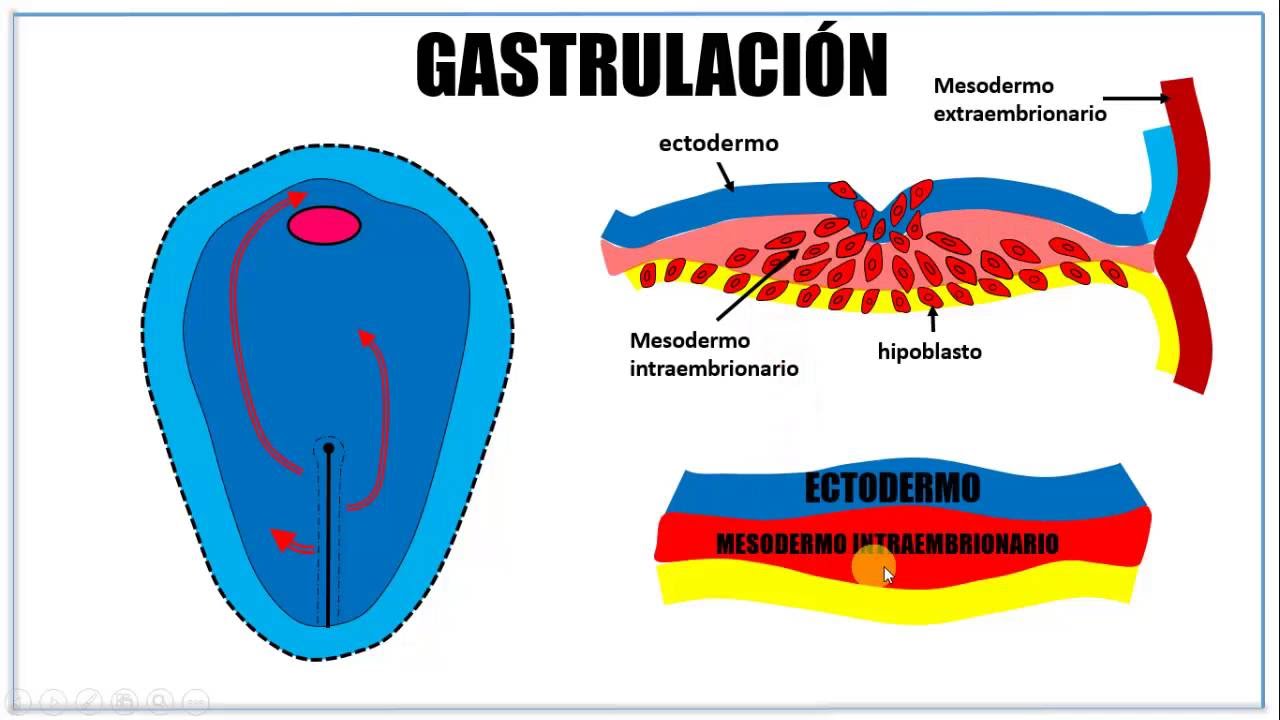
El mesodermo es una de las tres capas celulares embrionarias que surgen durante el proceso de gastrulación, alrededor de la tercera semana de gestación. Está presente en todos los vertebrados, incluyendo a los seres humanos.
Se define como una lámina blastodérmica que se localiza entre las capas ectodermo y endodermo. Antes de la gastrulación, el embrión sólo posee dos capas: el hipoblasto y el epiblasto.

Mientras que, durante la gastrulación, las células epiteliales de la capa epiblasto se convierten en células mesenquimatosas que pueden migrar a otras áreas. Dichas células se invaginan para dar lugar a las tres láminas o capas embrionarias.El mesodermo es la última capa que se origina, y se forma por un proceso de mitosis que ocurre en el ectodermo. Los animales que presentan esta capa reciben el nombre de “triblásticos” y entran dentro del grupo “bilateria”.Esta estructura se diferencia en tres áreas a cada lado de la notocorda: el mesodermo axial, el paraxial y el lateral. Cada una de estas partes va a dar lugar a diferentes estructuras del organismo.
Crea músculos y tejidos conjuntivos para todo el cuerpo, excepto en la parte de la cabeza, donde muchas estructuras proceden del ectodermo.
Por otro lado, posee la capacidad de inducir el crecimiento de otras estructuras como la placa neural, que es el precursor del sistema nervioso.
Desarrollo del mesodermo y sus derivados
El mesodermo da lugar principalmente a los músculos, huesos y vasos sanguíneos. Durante las primeras etapas del desarrollo embrionario, las células forman dos clases de tejidos:
Epitelios: las células se conectan a través de fuertes uniones construyendo láminas. El mesodermo forma numerosos epitelios.
Mesénquima: las células se distribuyen dejando amplios espacios entre ellas, constituyendo un tejido de relleno. La mesénquima es el tejido conectivo, y gran parte de ella proviene del mesodermo. Una pequeña parte surge del ectodermo.
Los derivados de esta estructura se explican mejor dividiéndolo en distintas áreas: mesodermo axial, paraxial y lateral. Ya que cada una de ella da lugar a estructuras distintas.

Mesodermo axial
Éste corresponde a una estructura fundamental en el desarrollo llamada notocorda. Ésta tiene forma de cordón, y se localiza en la línea media de la parte dorsal del embrión. Es el eje de referencia que determinará que ambos lados del cuerpo se desarrollen de forma simétrica.
La notocorda empieza a formarse a los 18 días de gestación, mediante unos movimientos celulares ocurridos durante el periodo de gastrulación. Comienza con una grieta superficial que se va plegando, y se invagina hasta formar un cilindro alargado.
Esta estructura es fundamental para determinar la posición del sistema nervioso y la posterior diferenciación neural. La notocorda tiene la importante función de desplegar señales inductivas que regulan el desarrollo del embrión.
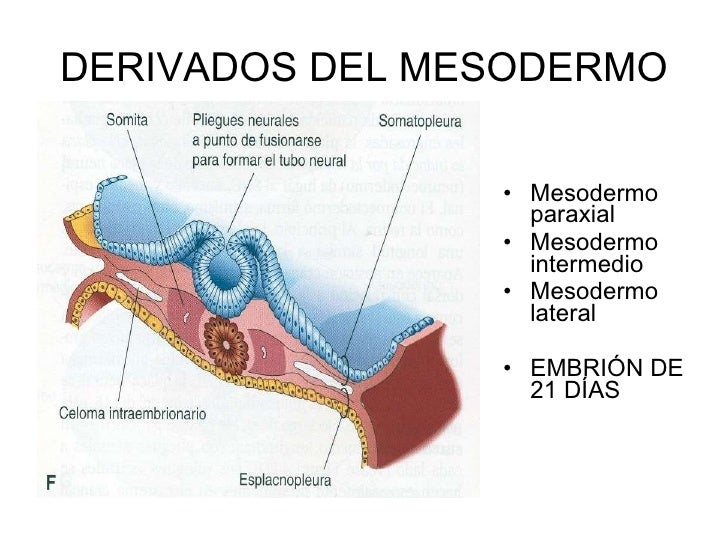
Mesodermo paraxial
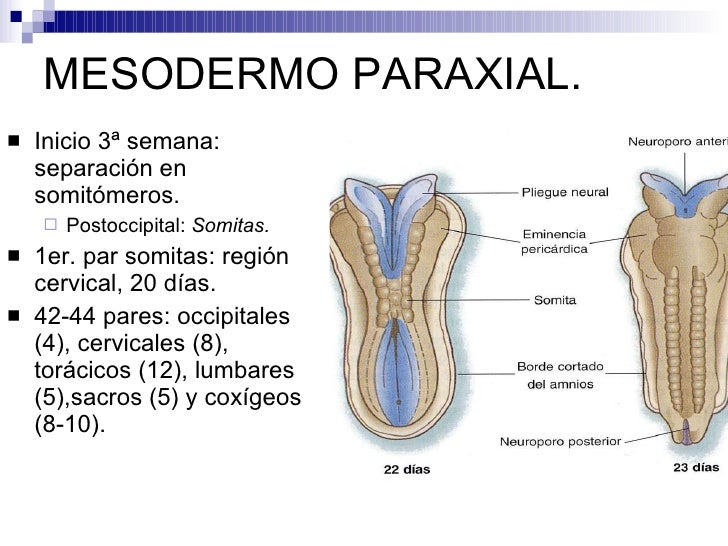
Es la parte más gruesa y ensanchada del mesodermo. Sobre la tercera semana, se divide en segmentos (llamados somitámeros) que van apareciendo en orden cefálico a caudal.
En el área cefálica, los segmentos se relacionan con la placa neuronal, formando neurómeras. Éstas van a dar lugar a gran parte del mesénquima cefálico.
Mientras que, en la zona occipital, los segmentos se organizan en somitas . Son estructuras transitorias fundamentales para la primera distribución segmentaria de la fase embrionaria temprana.
as somitas se disponen a ambos lados del tubo neural. Sobre la quinta semana, se observan 4 somitas occipitales, 8 cervicales, 12 torácicos, 5 lumbares, 5 sacros y 8-10 coccígeos. Éstos van a formar el esqueleto axial. Cada par de somitas va a evolucionar originando tres grupos de células:
– Esclerotoma: está formado por células que han migrado desde las somitas hasta la parte ventral de la notocorda. Éste va a convertirse en la columna vertebral, costillas, huesos del cráneo y cartílagos.
– Dermotoma: surge por las células de la parte más dorsal de los somitas. Da lugar al mesénquima del tejido conectivo, es decir, a la dermis de la piel. En aves, el dermotoma es el que produce la aparición de las plumas.
– Miotoma: da lugar a los músculos esqueléticos. Sus células precursoras son los mioblastos, que migran hacia la región ventral de los somitas.
Los músculos más cortos y profundos generalmente surgen de miotomos individuales. Mientras que superficiales y de gran tamaño, derivan de la fusión de varios miotomos. El proceso de formación de los músculos en el mesodermo se conoce como miogénesis.
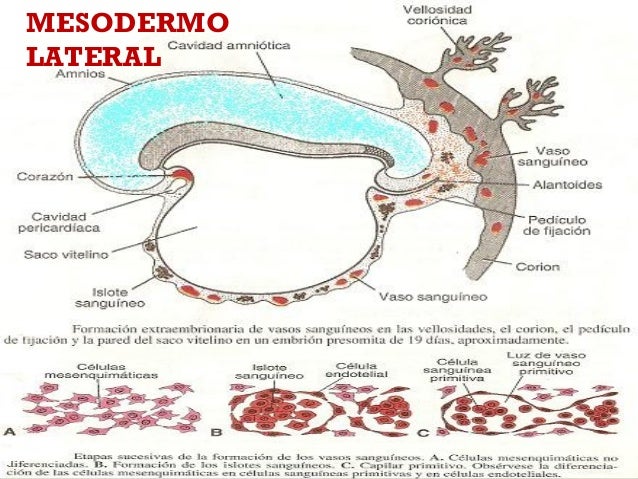
Mesodermo lateral
Es la parte más externa del mesodermo. A los 17 días de gestación aproximadamente, el mesodermo lateral se divide en dos láminas: el mesodermo esplacnopleural, que está al lado del endodermo; y el mesodermo somatopleural, que se localiza adyacente al ectodermo.
Por ejemplo, del mesodermo esplacnopleural provienen las paredes del tubo intestinal. Mientras que del mesodermo somatopleural surgen las membranas serosas que rodean las cavidades peritoneales, pleurales y pericárdicas.
Del mesodermo lateral surgen células que van a constituir el sistema cardiovascular y sanguíneo, el revestimiento de las cavidades corporales y la formación de membranas extraembrionarias. Éstas últimas tienen la misión de llevar nutrientes al embrión.
En concreto, da lugar al corazón, vasos sanguíneos, células sanguíneas como glóbulos rojos y blancos, etc.
VIDEO COMPLEMENTARIO:
https://www.youtube.com/watch?v=jHafs96_wRs
ENDODERMO:
El endodermo es una de las tres capas germinales que surgen en el desarrollo embrionario temprano, sobre la tercera semana de gestación. Las otras dos capas se conocen como ectodermo o capa externa y mesodermo o capa media. Debajo de éstas se encontraría el endodermo o capa interna, que es la más fina de todas.
Antes de la formación de estas capas, el embrión está compuesto por una única lámina de células. A través del proceso de gastrulación, el embrión se invagina (pliega sobre sí mismo) para producir las tres capas de células primitivas. Primero aparece el ectodermo, luego el endodermo y finalmente, el mesodermo.

Antes de la gastrulación, el embrión es sólo una capa de células que luego se divide en dos: el hipoblasto y el epiblasto. Sobre el día 16 de gestación, una serie de células migratorias fluyen por la raya primitiva, desplazando a las células del hipoblasto para transformarse en el endodermo definitivo.
Más adelante, se produce un fenómeno llamado organogénesis. Gracias a éste, las capas embrionarias comienzan a cambiar para transformarse en los diferentes órganos y tejidos del organismo. Cada capa va a dar lugar a diferentes estructuras.
En este caso, el endodermo, va a originar el sistema digestivo y respiratorio. También forma el revestimiento epitelial de muchas partes del cuerpo.
Al principio el endodermo está formado por células aplanadas, que son células endoteliales que forman tejidos de revestimiento principalmente. Son más anchas que altas. Más adelante se convierten en células columnares, que significa que son más altas que anchas.
Una de las capas más antiguas de la diferenciación embrionaria en los seres vivos es el endodermo. Por ese motivo, provienen de ella los órganos más importantes para la supervivencia del individuo.
Desarrollo del endodermo
La diferenciación del cuerpo del embrión del líquido exterior afecta al endodermo, dividiéndolo en dos partes: el endodermo embrionario y el extraembrionario.
No obstante, los dos compartimentos se comunican por una apertura amplia, precursora del cordón umbilical.
Endodermo embrionario
Es la parte del endodermo que va a formar estructuras dentro del embrión. Da lugar al intestino primitivo.
Esta capa germinal es la responsable, junto con el mesodermo, de originar la notocorda. La notocorda es una estructura que tiene importantes funciones. Una vez formada, se sitúa en el mesodermo, y se encarga de transmitir señales inductivas para que las células migren, se acumulen y se diferencien.
La transformación del endodermo va paralela a los cambios inducidos por la notocorda. Así, la notocorda induce pliegues que van a determinar los ejes craneal, caudal y lateral del embrión. El endodermo también se pliega progresivamente hacia el interior de la cavidad corporal por la influencia de la notocorda.
Al principio comienza con el llamado surco intestinal, que se invagina hasta cerrarse y formar un cilindro: el tubo intestinal.
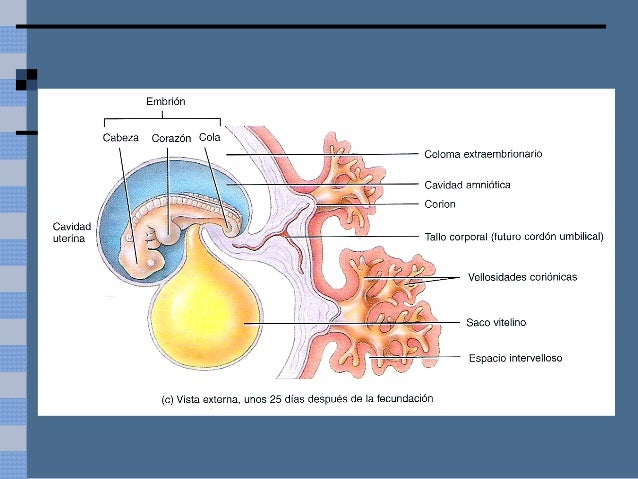
Endodermo extraembrionario
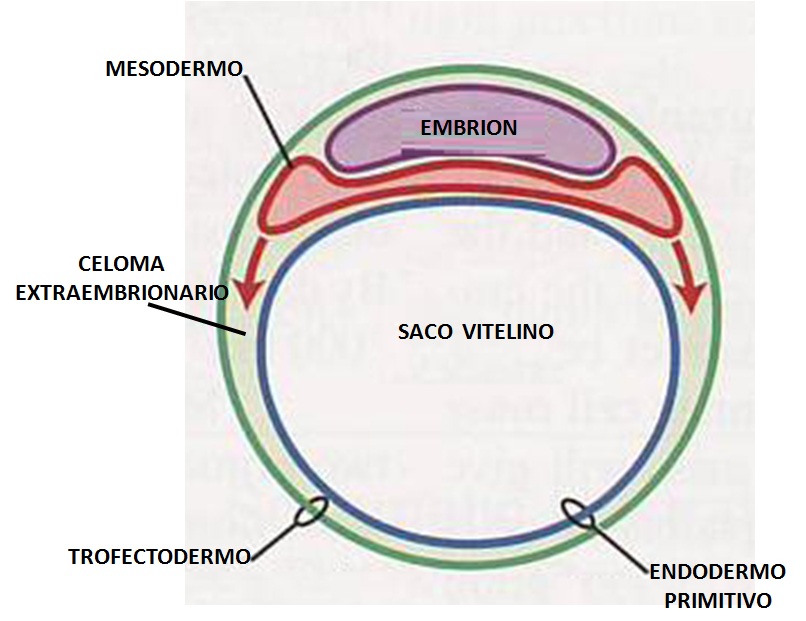
La otra porción de endodermo queda fuera del embrión, y se denomina saco vitelino. El saco vitelino consiste en una estructura membranosa adherida al embrión que se encarga de nutrir, dar oxígeno a éste y eliminar desechos.
Sólo existe en etapas tempranas del desarrollo, hasta la décima semana de gestación aproximadamente. En los seres humanos, este saco ejerce la función del sistema circulatorio.
Partes del tubo intestinal del endodermo
Por otro lado, se pueden diferenciar distintas áreas en el tubo intestinal del endodermo. Cabe decir que algunas de ellas pertenecen al endodermo embrionario y otras al extraembrionario:
– El intestino craneal o interior, que se encuentra dentro del pliegue de la cabeza del embrión. Comienza en la membrana bucofaríngea, y esta región va a convertirse en la faringe. Luego, en el extremo inferior de la faringe aparece una estructura que originará el tracto respiratorio.
Debajo de esta área, el tubo se ensanchará rápidamente para convertirse posteriormente en el estómago.
– Intestino medio, situado entre el intestino craneal y caudal. Éste se prolonga hasta el saco vitelino mediante el cordón umbilical. Esto permite al embrión recibir nutrientes del organismo de su madre.
– El intestino caudal, dentro del pliegue caudal. A partir de él surge el alatoides, una membrana extraembrionaria que aparece por una invaginación localizada al lado del saco vitelino.

Derivados del endodermo
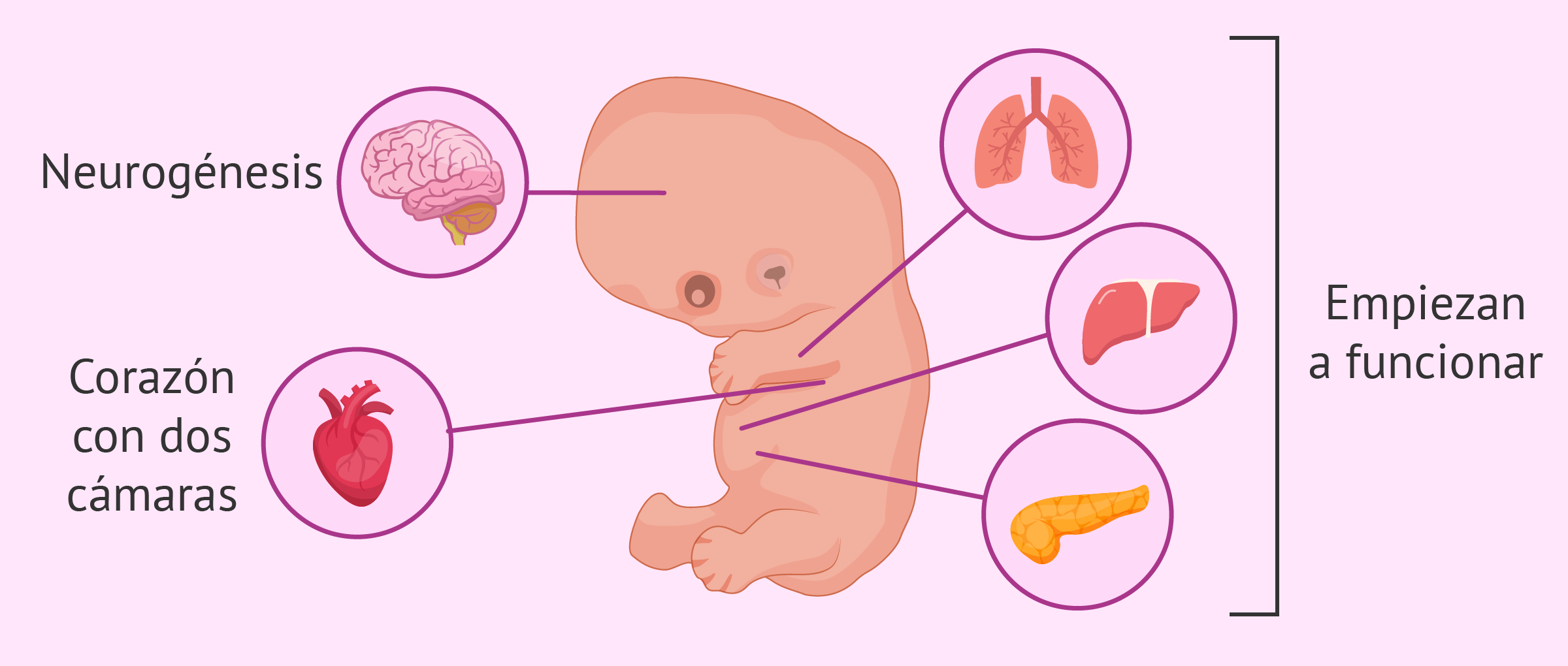
Como se mencionó, el endodermo deriva en órganos y estructuras del cuerpo a través de un proceso llamado organogénesis. La organogénesis ocurre en una etapa que dura desde la tercera a la octava semana de gestación aproximadamente.
El endodermo contribuye en la formación de las siguientes estructuras:
– Glándulas del tracto gastrointestinal y órganos gastrointestinales asociados como el hígado, la vesícula biliar y el páncreas.
– Epitelio o tejido conectivo que rodea: las amígdalas, la faringe, la laringe, la tráquea, los pulmones, y el tracto gastrointestinal (menos la boca, el ano, y parte de la faringe y el recto; que provienen del ectodermo).
También forma el epitelio de la trompa de Eustaquio y la cavidad timpánica (en el oído), las glándulas tiroideas y paratiroideas, la glándula timo, la vagina y la uretra.
– Tracto respiratorio: como bronquios y alvéolos pulmonares.
– Vejiga urinaria.
– Saco vitelino.
– Alantoides.
Se ha visto que en los seres humanos el endodermo puede diferenciarse en órganos observables tras las 5 semanas de gestación.

TEJIDO MUSCULAR:
El tejido muscular se forma por una derivación mesodérmico responsable del movimiento de los órganos que son de las partes del cuerpo de un ser vivo que desempeñan una función diferenciada de los organismos que está formado por unas células muy alargadas denominadas fibras musculares o miocitos que es una célula fusiforme que tiene forma de huso y multinucleada son células eucariotas que tienen más de un núcleo en su interior con capacidad de contraerse con mucha facilidad, además cada una de las fibras musculares están envuelta por una fina lámina de tejido conjuntivo o conectivo que es un conjunto heterogéneo de tejidos orgánicos que comparten un origen común a partir del mesénquima embrionario originado a partir del mesodermo.
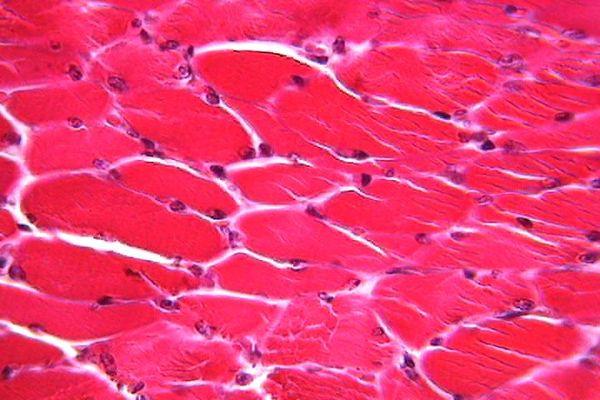
El tejido muscular liso, es el que está formado por la unión de varias células largas que están preparadas en capas más que todo en las paredes de los órganos huecos, como el tubo digestivo o vasos sanguíneos. pero de esta manera encuentran células musculares lisas en el tejido conjuntivo que afecta ciertos órganos como la próstata y las vesículas seminales y en el tejido subcutáneo que es cuando se produce en determinadas regiones como el escroto que son bolsa que están formada por la piel que cubre los testículos de los mamíferos y los pezones.

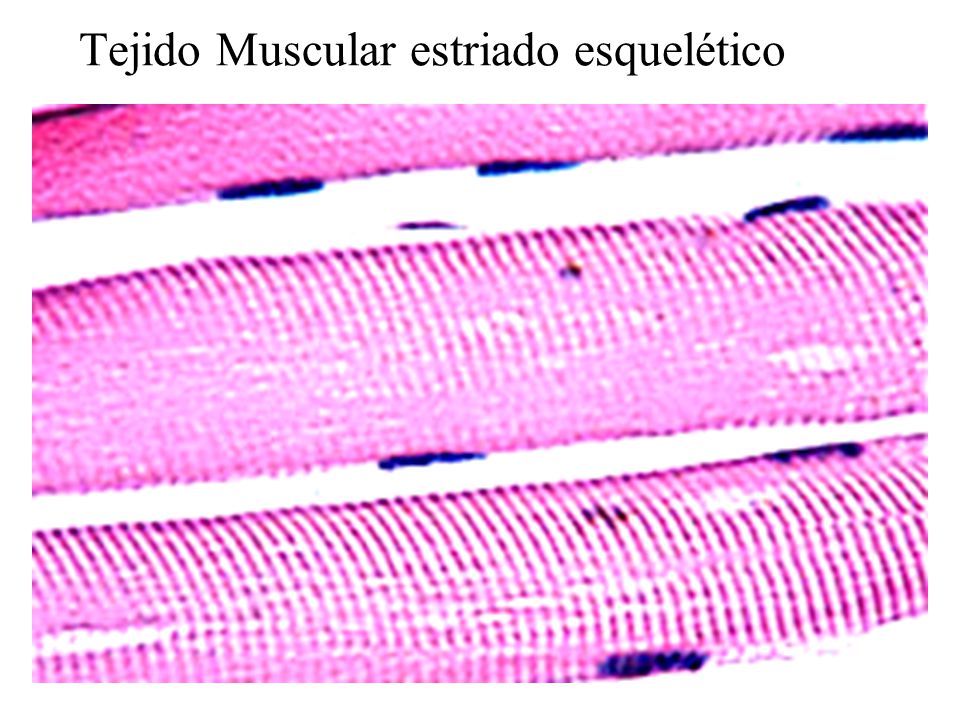
El tejido muscular estriado o esquelético, son los que están formados por las células muy largas cubierta por una membrana externa de tejido conjuntivo denominado empimisio. Además de están parten septos que son tabique que se divide de un modo completo o incompleto de una cavidad o partes del cuerpo en un animal que se dirigen hacia el interior del músculo.

El tejido muscular cardíaco, son las que están constituidas por células alargadas que también presentan estriaciones transversales de la disposición de las proteínas contráctiles y pueden distinguirse fácilmente de las fibras musculares esqueléticas por el hecho de poseer uno o dos núcleos centrales.


TEJIDO NERVIOSO:
El tejido nervioso es el conjunto de células especializadas que forman el sistema nervioso. Las funciones más importantes del tejido nervioso son recibir, analizar, generar, transmitir y almacenar información proveniente tanto del interior del organismo como fuera de éste. Es un complejo sistema encargado de regulación de diversas funciones orgánicas vitales como son la respiración, la alimentación, la digestión, el sueño, etc. También es el origen de funciones muy complejas y abstractas como el pensamiento, la memoria y el aprendizaje.
Desde el punto de vista anatómico, el sistema nervioso puede dividirse en sistema nervioso central (SNC) —que incluye el encéfalo y la médula espinal— y sistema nervioso periférico (SNP) —el cual comprende los nervios espinales, los nervios craneales y sus ganglios relacionados—. Desde un punto de vista funcional también se puede dividir en sistema nervioso somático o voluntario y sistema nervioso autónomo (que, a su vez, se subdivide en sistema simpático y parasimpático).


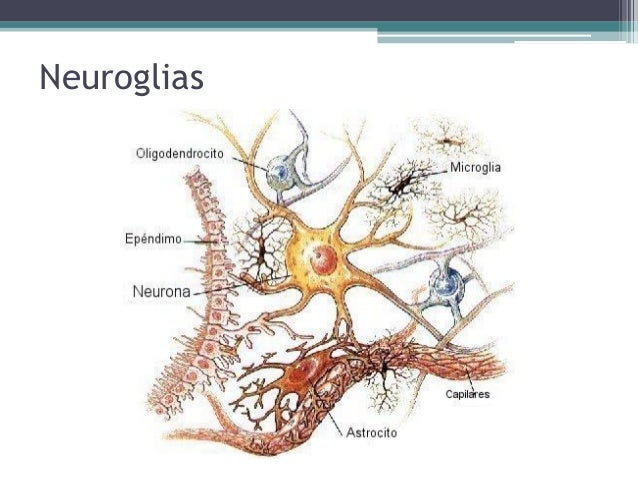
El tejido nervioso está constituido por dos tipos de células: 1) las neuronas, su función está basada en el desarrollo de dos propiedades que son la excitabilidad y la conductividad; las neuronas son las encargadas de recibir estímulos del medio, transformarlos e integrarlos, así como transmitirlos como impulsos, integradores cognitivos y motores del sistema nervioso. 2) Las células de la glía o neuroglía, encargadas de desempeñar diversas funciones: de soporte, defensa, mielinización, nutrición a las neuronas, regulación de la composición del microambiente, protección, formar parte de la barrera hematoencefálica, revestimiento, formación de líquido cefalorraquídeo, reparación de daño cerebral, fagocitosis, etcétera.
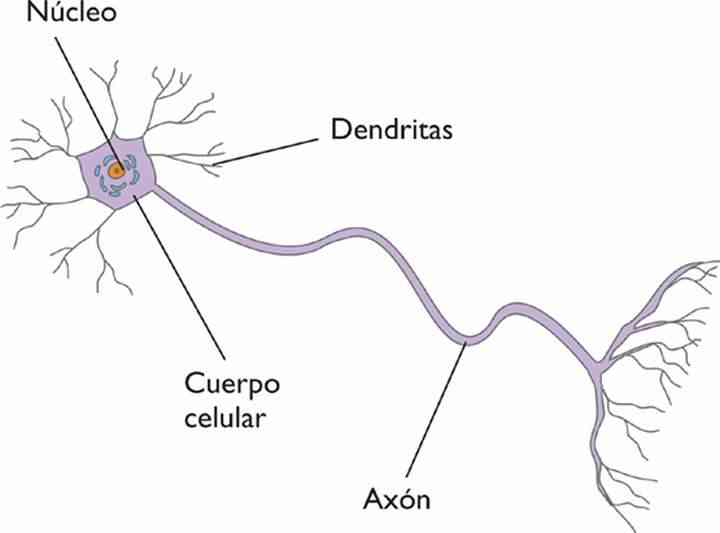
Neurona
La neurona es el elemento principal en el funcionamiento del tejido nervioso, son células especializadas en recibir señales desde receptores sensoriales, que conducen y transmiten impulsos eléctricos que consisten en cambios en la polaridad eléctrica a nivel de su membrana celular; este grado de especialización conlleva, entre otras cosas, a la nula capacidad de división.
La forma y estructura de cada neurona se relaciona con su función específica, la que puede seR:
- recibir señales desde receptores sensoriales
- conducir estas señales como impulsos nerviosos, que consisten en cambios en la polaridad eléctrica a nivel de su membrana celular
- transmitir las señales a otras neuronas o a células efectoraS
ganglios del SNP (Fig 7 y 8)
Los axones forman la parte funcional de las fibras nerviosas y se concentran en los haces de la sustancia blanca del SNC; y en los nervios del SNP
Tipos de neuronas
  | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Figura 1 Figura 2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Según el número y la distribución de sus prolongaciones, las neuronas se clasifican en:
Uniones intercelulares de las neuronas
Las neuronas se asocian entre sí y con otras células en estructuras especializadas que permiten:
Transporte axónico
El transporte de organelos, enzimas, agregados macromoleculares y metabolitos, es una función de axoplasma en la cuál intervienen directamente los microtúbulos (Fig 1).
Ocurre en dos direcciones:
La velocidad del transporte varía entre:
Los transportes axonales rápidos están mediados por la interacción molecular entre microtúbulos y los dos moléculas son capaces de desplazarse a lo largo de los microtúbulos.
En el flujo rápido anterógrado es la molécula de kinesina, unida a un receptor en la membrana del organelo transportado que se desplaza, a expensas de ATP, desde el extremo (-) del microtúbulo, ubicado en el pericarion hacia su extremo (+)
En el flujo rápido retrógrado es la molécula de dineína citoplasmática (MAP1C) unida a un receptor en la membrana del organelo transportado la que se desplaza interactuando con la tubulina a expensas de ATP, desde el extremo (+) del microtúbulo, ubicado en el terminal axónico hacia su extremo (-)
Sinapsis
Conducen el impulso nervioso sólo en una dirección. Desde el terminal pre-sináptico se envián señales que deben ser captadas por el terminal post-sináptico.
Existen dos tipos de sinapsis, eléctricas y químicas que difieren en su estructura y en la forma en que transmiten el impulso nervioso.
Sinapsis eléctricas: corresponden a uniones de comunicación entre las membranas plasmáticas de los terminales presináptico y postsinápticos . las que al adoptar la configuración abierta permiten el libre flujo de iones desde el citoplasma del terminal presinático hacia el citoplasma del terminal postsináptico..
Sinapsis química: se caracterizan porque las membranas de los terminales presináptico y postsináptico están engrosadas y las separada la hendidura sinátpica, espacio intercelular de 20-30 nm de ancho. El terminal presináptico se caracteriza por contener mitocondrias y abundantes vesículas sinápticas, que son organelos revestidos de membrana que contienen neurotransmisores (Fig 1)
Al fusionarse las vesículas sinápticas con la membrana se libera el neurotrasmisor que se une a receptores específicos localizados en la membrana post-sináptica, en la cuál se concentran canales para cationes activados por ligandos
Al llegar el impulso nervioso al terminal presináptico se induce: la apertura de los canales para calcio sensibles a voltaje, el subir el ncalcio intracelular se activa la exocitosis de las vesículas sinápticas que liberan al neurotransmisor hacia la hendidura sináptica. La unión del neurotrasmisor con su receptor induce en la membrana postsinática la apertura de los canales para cationes activados por ligandos determinando cambios en la permeabilidad de la membrana que pueden: inducir la depolarización de la membrana postsinática: sinápsis exhitatorias; o hiperpolarizar a la membrana postsináticas: sinapsis inhibitorias.
La sumatoria de los impulsos exitatorios e inhibitorios que llegan por todas las sinapsis que se relacionan con a cada neurona( 1000 a 200.000) determina si se produce o no la descarga del potencial de acción por el axón de esa neurona.
Células de sostén
En el tejido nervioso del SNC, por cada neurona hay entre 10 a 50 células de neuroglia (Fig 1), y que a diferencia de las neuronas retienen su capacidad de proliferar
Existen 4 clases de células de neuroglia:
En el tejido nervioso del SNP, tanto las neuronas, en los ganglios, como los axones ubicados en las fibras nerviosas, están rodeadas por de células de sostén (Fig. 2).
Se distinguen dos tipos:
Astrocitos
Tienen formas estrelladas y presentan largas prolongaciones que se extienden hacia las neuronas y hacia los láminas basales que rodean a los capilares sanguíneos (pies terminales) (Fig 1), o que separan al tejido nervioso del conjuntivo laxo de la piamadre, constituyendo la glia limitante
Las prolongaciones de los astrocitos contienen manojos de filamentos intermedios específicos formados por la proteína ácida fibrilar.
Se han identificado dos tipos de astroglia: astrocitos fibrosos que se asocian de preferencia a las fibras nerviosas de la sustancia blanca (Fig 2). y astrocitos protoplasmáticos que se concentran de preferencia asociados a los pericariones, dendritas, terminaciones axónicas en la sustancia gris (Fig 3)
Oligodendrocitos
Son más pequeños y con menos prolongaciones que la astroglía (Fig. 1).
Su núcleo es rico en heterocromatina y su citoplasma contiene ergastoplasma, polirribosomas libres, un aparato de Golgi desarrollado y un alto contenido n microtúbulos, tanto en el citoplasma que rodea al núcleo como en sus prolongaciones. Su función más notable es la formación de la mielina, que rodea a los axones del SNC.
El proceso de mielinización del axón por el oligodendrocito es similar al de la célula de Schwann. Sin embargo una oligodendroglia puede formar mielina en cada una de sus prolongaciones que se adhieren inicialmente a un axón, de modo que internodos mielinizados de varios axones dependen un oligodendrocito.
Células ependimarias
Forman un tipo de epitelio monostratificado que reviste las cavidades internas del SNC que contienen al líquido cefalo raquídeo (ventrículos y conducto del epéndimo).
Se unen entre sí por comlejosde unión similares a los epiteliales pero carecen de zona de oclusión, de modo que el liquido cefaloraquídeo se comunica con los espacios intercelulares existentes entre las células nerviosas y las glías. Presentan además largas prolongaciones en su zona basal que se asocian a las prolongaciones de la astroglia y en su superficie apical presenta microvellocidades y cilios.
Microglia
Se caracterizan por ser pequeñas, con un denso núcleo alargado y prolongaciones largas y ramificadas.
Contienen lisosomas y cuerpos residuales. Si bien se la clasifica generalmente como célula de la neuroglia ellas presentarnel antígeno común leucocítico y el antígeno de histocompatibilidad clase II, propio de las células presentadoras de antígeno.
Células de Schwann
Las células de Schawnn se originan de la cresta neural y acompañan a los axones durante su crecimiento, formando la vaina que cubre a todos los axones del SNP desde su segmento inicial hasta sus terminaciones. Ellas son indispensables para la integridad estructural y funcional del axón.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Son células pequeñas localizadas en los ganglios, dendritas y terminales axónicos.Están rodeadas por lámina basal y separan a las células nerviosas del estroma fibrocolagenoso presente en lel tejido propio del SNP.
Estructura básica del SNP
En el SNP las células nerviosas se organizan formando dos tipos de estructuras:
TEJIDO CONJUNTIVO:
Los tejidos conectivos reúnen, dan apoyo y protegen a los otros tipos de tejido.
Está conformado por:
Función: Sostén, relleno, defensa y nutrición.
Las cápsulas que revisten los órganos y la malla tridimensional interna que soporta sus células están constituidas por tejido conjuntivo. Así también los tendones, ligamentos y el tejido aerolar que llena los espacios entre los órganos.
Esta íntimamente asociado con los vasos sanguíneos, está directamente involucrado en la nutrición. Tanto las sustancias nutritivas transportadas por la sangre como los productos de desecho del metabolismo que son conducidos a los órganos de eliminación atraviesan el conjuntivo que envuelve a los capilares.
FIBRAS: se distribuyen desigualmente en las variedades de tejido conjuntivo. Pueden existir más de un tipo de fibras en un mismo tejido. Las fibras predominantes son las responsables de ciertas propiedades del tejido.
 SE CLASIFICA EN:
Tejido conjuntivo laxo
Se llama también areolar, y es tal vez el tejido conjuntivo más común. Rellena los espacios entre las fibras y haces musculares, sirve de apoyo para los epitelios y forma una capa alrededor de los vasos sanguíneos y linfáticos. Apoyando y nutriendo las células epiteliales, el tejido conjuntivo laxo se encuentra en la piel, en las mucosas y en las glándulas.
El tejido conjuntivo laxo contiene todos los elementos del conjuntivo propiamente dicho. No hay predominio absoluto de ninguno de los componentes. Las células más comunes son los fibroblastos y los macrófagos, aun cuando los otros tipos descritos están presentes las fibras colágenas, elásticas y reticulares también están presentes.
El tejido conjuntivo laxo es de consistencia delicada, flexible y poco resistente a las tracciones.

Tejido conjuntivo denso
Esta variedad está formada por los mismo elementos estructurales hallados en el tejido conjuntivo laxo, con predominancia acentuada de las fibras colágenas. En los cortes se observa que las células son menos numerosas que en el conjuntivo laxo y que entre ellas sobresalen los fibroblastos. Se trata de un tejido menos flexible que el laxo y mucho más resistente a las tracciones. Cuando las fibras colágenas se disponen en haces sin orientación tenemos el tejido denso no modelado (irregular). En este tejido los haces forman una trama tridimensional, lo que da al tejido cierta resistencia a las tracciones ejercidas en cualquier dirección. el tejido conjuntivo denso no modelado se encuentra por ejemplo en la dermis profunda de la piel.
El tejido conjuntivo denso modelado presenta los haces colágenos orientados según una organización fija. Se trata de un conjuntivo que formó sus fibras colágenas como respuesta a tracciones ejercidas en determinado sentido. Las fibras se orientan de modo que ofrezcan el máximo de resistencia a las fuerzas que normalmente actúan sobre el tejido. Los tendones presentan el más típico ejemplo de tejido denso modelado.
Los tendones son estructuras cilíndricas alargadas que unen los músculos esqueléticos a los huesos. Debido a su riqueza en fibras colágenas, son blancos inextensibles. Están formados por haces paralelos de fibras colágenas, entre los cuales existe poca cantidad de sustancia fundamental amorfa y de fibroblastos con ciertas características propias. Estos fibroblastos tienen núcleos alargados paralelos a las fibras y citoplasma delgado que tiende a envolver los haces de fibras de colágena.
Los haces de fibras colágenas del tendón (haces primarios) forman conjuntos (haces secundarios) envueltos por el tejido conjuntivo laxo que contiene vasos sanguíneos y nervios. Finalmente, el tendón esta envuelto externamente por una vaina de tejido conjuntivo denso. en algunos tendones esta vaina está dividida en dos capas: una sujeta al tendón y otra unida a las estructuras vecinas. Se forma así una cavidad revestida de células aplanadas y de origen mesenquimatoso, en la que hay líquido semejante al sinovial de las articulaciones, que contienen agua, glucosaminoglicanos, proteínas e iones y que facilita el deslizamiento del tendón.

Tejido elástico
Este tejido está formado por fibras elásticas gruesas paralelas y organizadas en haces separados por tejido conjuntivo laxo. Entre las fibras elásticas se observan fibroblastos aplanados, como se encuentran en los tendones. La riqueza en fibras elásticas confiere al tejido un color amarillo típico y de gran elasticidad. El tejido elástico es poco frecuente, encontrándose por ejemplo, en los ligamentos amarillos de la columna vertebral y el ligamento suspensor del pene.
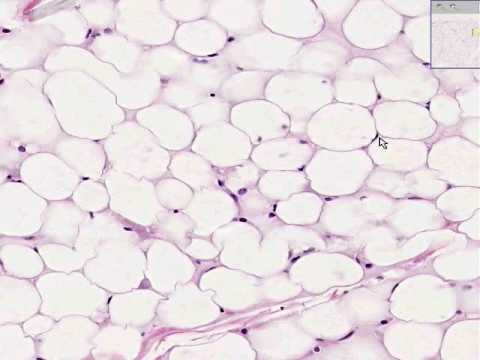
 Tejido adiposo
El tejido adiposo es un tipo especial de tejido conjuntivo, en que se observa gran predominio de células adiposas, que se caracterizan por almacenar grasa neutras. Estas células pueden hallarse aisladas o en pequeños grupos en el tejido conjuntivo común pero la mayoría de ellas se agrupan en el tejido adiposo distribuido por el cuerpo.
Como los mamíferos consumen energía de modo continuo, pero se alimentan intermitentemente, se comprende la importancia de un reservorio de energía representado por el tejido adiposo. Además de esta función el tejido adiposo tiene otras; por localizarse debajo de la piel modela la superficie corporal. Forma también almohadillas amortiguadoras como en la planta de los pies y palma de las manos. Como la grasa son malas conductoras de calor, este tejido contribuye al aislamiento térmico del individuo. Además llena espacios entre los tejidos y ayuda a mantener a ciertos órganos en su posición normal. En animales que hibernan hay una variedad particular de tejido adiposo que suministra el calor necesario para que el animal pueda volver a la actividad, después del período de hibernación.
Hay dos variedades de tejido adiposo identificado por la estructura de sus células y por su localización, color, inervación, vascularización y funciones. Una variedad es el tejido adiposo común, amarillo o unilocular, cuyas células, cuando están completamente desarrolladas, contienen sólo una gota de grasa en el citoplasma. La otra variedad es el tejido adiposos pardo, formado por células que contienen numerosas gotitas de lípidos, siendo conocido también como tejido adiposo multilocular.
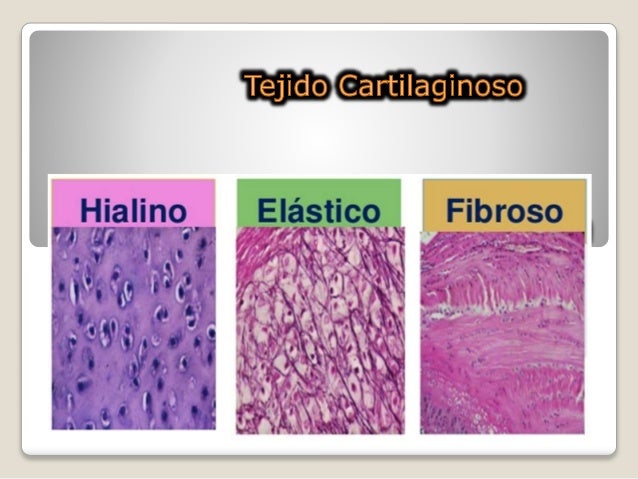
 Tejido cartilaginoso
El cartílago tiene una consistencia rígida, menos consistente que el tejido óseo. Su superficie el ligeramente elástica y lisa facilitando los desplazamientos. Desempeña la función de soporte a la cual se suma la de revestir superficies articulares facilitando los movimientos. Al igual que los demás tipos del conjuntivo, el tejido cartilaginoso contiene células, lo condrocitos, y abundante material intercelular que forma la matriz. Representa uno de los primeros tejidos adaptados para soportar peso.
Las propiedades del tejido cartilaginoso relacionadas con su papel fisiológico dependen de la estructura fisicoquímica de la matriz, está constituida por colágena, esta más elastina, en asociación con macromoléculas de glucosaminoglicanos. La función de las células consiste en producir una matriz con características adecuadas y mantenerla en estado normal. La muerte de las células conlleva a la generación de la matriz. Los glucosaminoglicanos se unen químicamente a las proteínas formando proteoglicanos de elevado peso molecular y de gran viscosidad. Los proteoglicanos se asocian por medio de uniones químicas débiles a colágena de la matriz. Los proteoglicanos confieren rigidez al cartílago porque las fibras elásticas y de colágena son flexibles.
Las variaciones en la cantidad y tipo de las fibrillas dan propiedades esenciales al cartílago. En las áreas sujetas a agrandes o tracciones, la cantidad de fibras de colágena es más elevada, lo que hace que el cartílago sea prácticamente inextensible y muy resistente a la tracción. La presencia de fibras elásticas, con reducción del porcentaje de colágena, da origen a un tipo de cartílago que es poco rígido, flexible y elástico.
El tejido cartilaginoso no posee vasos sanguíneos, siendo nutrido por los capilares del conjuntivo que los rodea o a través del líquido sinovial de las cavidades articulares. El algunos casos los vasos sanguíneos atraviesan los cartílagos, yendo a nutrir otros tejidos. El cartílago esta desprovisto de vasos linfáticos y nervios. Tiene un metabolismo bajo.
Hay tres tipos de tejido cartilaginoso, clasificados de acuerdo a la abundancia y al tipo de fibra presente en la matriz: 1) cartílago hialino que es el más común y cuya matriz posee una cantidad moderada de fibras colágena; 2) cartílago elástico, que posee fibras de colágena y abundantes fibras de elastina y 3) cartílago fibroso, que presenta la matriz constituida casi completamente de fibras de colágena.
Los cartílagos (excepto los articulares y las piezas de cartílago fibroso) están envueltos por una vaina conjuntiva que recibe el nombre de pericondrio, el cual se prolonga gradualmente con el cartílago por un lado y con el conjuntivo adyacente por el otro.
 Tejido óseo
Es uno de los más resistentes y rígidos del cuerpo humano. Como tejido especializado en soportar presiones, sigue el cartílago, tanto en la ontogénesis como en la filogénesis. Como constituyente principal del esqueleto sirve de soporte para las partes blandas y protege órganos vitales, como los contenidos en las cajas craneana y torácica y el conducto raquídeo.
Aloja y protege la médula ósea, formadora de las células de la sangre. Además de estas funciones, proporciona apoyo a los músculos esqueléticos transformando sus contracciones en movimientos útiles y constituye un sistema de palancas que incrementa las fuerzas generadas por la con tracción muscular.
El tejido óseo esta formado por células y un material extracelular calcificado, la matriz ósea . Las estructuras son: 1) Los osteocitos que se sitúan en las cavidades o algunas en el interior de la matriz; 2) los osteoblastos, productores de la parte orgánica de la matriz, y 3) los osteoclastos, células gigantes multinucleadas, relacionadas con la resorción del tejido óseo, que participan en los procesos de remodelación de los huesos.
Como no existe difusión de sustancias a través de la matriz calcificada del hueso, la nutrición de los osteocitos depende de canalículos que existen en la matriz. Esos canalículos permiten la comunicación de los osteocitos con sus vecinos, con las superficies externa e interna del hueso y con los canales vasculares de la matriz.
Todos los huesos están revestidos en sus superficies externa e internas por membranas conjuntivas, el periostio y el endostio, respectivamente.
 Tejido Sanguineo: El tejido sanguíneo se caracteriza porque está constituido por células libres que son los eritrocitos, los leucocitos y plaquetas llamados en conjunto elementos figurados de la sangre y por su matriz extracelular líquida conocida como plasma sanguíneo. Las preparaciones para estudiar el tejido sanguíneo deben ser frotis del líquido, es decir, la toma de la muestra se hace por punción para obtener una gota de sangre, la cual se deposita sobre un portaobjetos limpio y desgrasado, luego se extiende con el borde de otro portaobjetos para formar una capa delgada, se seca y se tiñe con solución de Wrigth (mezcla de colorantes ácidos y básicos) para contrastar e identificar los diversos elementos celulares.  |






































No hay comentarios:
Publicar un comentario